


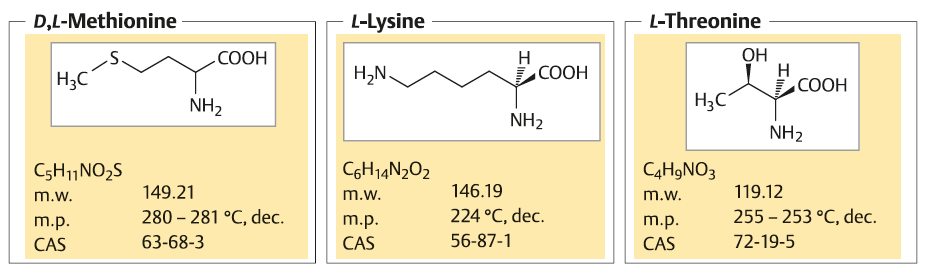
کلیات D-L-Methionine, L-Lysine, and L-Threonine :
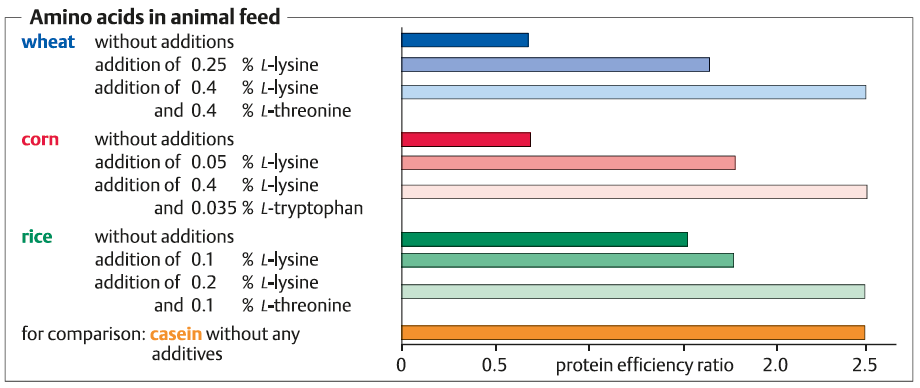
این سه اسید امینه عمدتا به عنوان افزودنی های خوراکی استفاده می شود . برای انسان ها و اکثر حیوانات اهلی , اسید آمینه های ضروری هستند که در بدن موجودات تولید نمی شوند . محصولات کشاورزی زیادی که به عنوان خوراک استفاده می شوند مانند ذرت , سویا , جو , گندم , برنج مقدار کافی از این اسید های آمینه را که در یک تغذیه سالم لازم است ندارند .به همین دلیل توصیه می شود که رژیم گیاهخواری با این اسید های امینه تکمیل شود .
در خوراکی ها این کمبود نقش مهم تری را ایفا می کند .از انجاییکه ک افزایش وزن طی چاقی یک جاندار تغذیه کننده از گندم یا برنج تنها زمانی به استاندارد تغذیه ای کازئین دست پیدا میکنند که L- لیزین و L- ترئونین افزوده شود . به همین ترتیب , یک تغذیه برمبنای محصولات کشاورزی نیاز به اضافه شدن L و D متیونین، L-لیزین، L-تریپتوفان دارد . این اسید های امینه به صورت صنعتی طی سنتز شیمیایی یا تخمیر تولید می شوند.
D-L-متیونین :
سنتز این اسید امینه طی پنج مرحله طی می باشد که شامل آکرولین ( acrolein ) , متان اتیول (methanethiol ) و HCN که به سمت تشکیل واسطه از یک هیدانتویین (hydantoin )هدایت می شوند و از آنجاییکه D-متیونین در جانداران عالی به L-متیونین تبدیل می شود تجمع D-L-متیونین میتواند به عنوان یک افزودنی خوراکی استفاده شود . جداسازی انانتیومر ها ضروری نیست ; مقدار کمی از L-متیونین توسط هیدولیز مشتقات D-L-متیونین به واسطه آنزیم هایی که به صورت انتخابی مشتق L-انانتیومر را جداسازی می کنند ساخته می شوند .
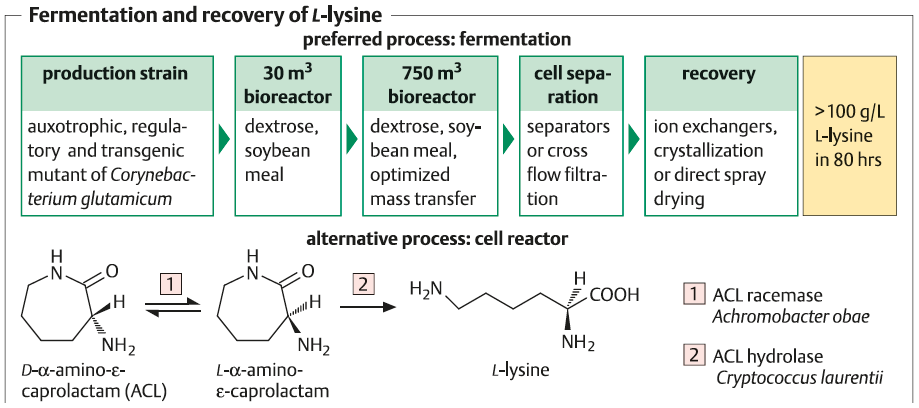
L-لیزین :
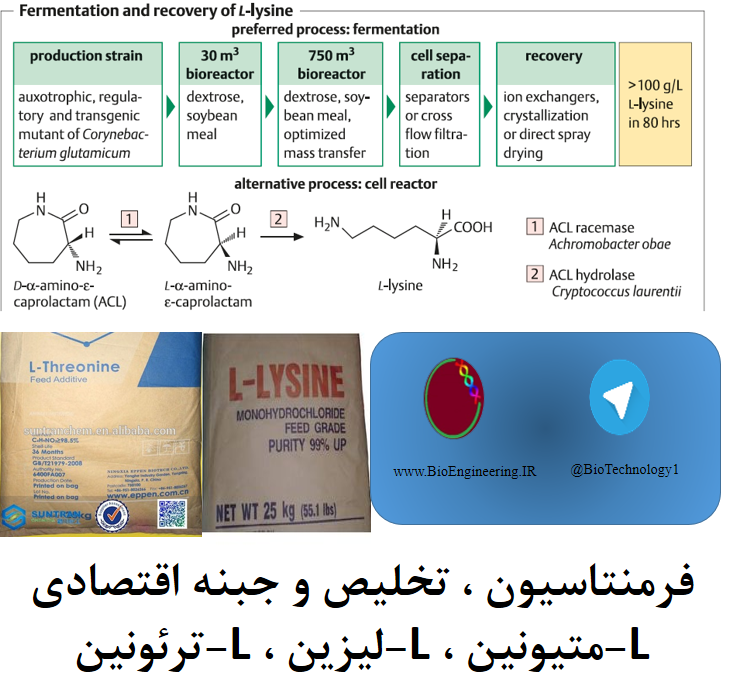
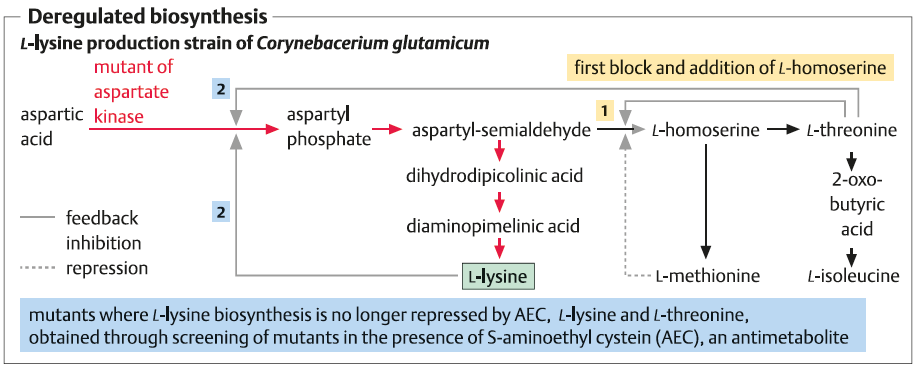
عمدتا توسط تخمیر با استفاده از باکتری جهش یافته Corynebacterium glutamicum تولید می شود . که به واسطه اگزالواستات در چرخه اسیدسیتریک بین محل تجمع آسپارتیک اسید و اسید پیروات در یک مسیر چند مرحله ای تولید کننده دی آمینو پالمیلیک اسید (DAP) که به عنوان حد واسط است تهیه می شوند . مسیر های جانبی این چرخه به سمت تولید L-ترئونین و L-متیونین پیش می روند که میتوانند تولید لیزین را با مهار بازخورد آن متوقف کنند .
در سویه های جهش یافته بیش تولید لیزین , این فرآیند به علت رفع محدودیت یا دور زدن انزیم های قوی تنظیم کننده از طریق جهش های اگزوتروفیک (auxotrophic) حذف می شوند .جهش های مقاوم S-آمینواتیل سیستئین AEC نقش ویژه ای را ایفا میکند , در نتیجه جهش های مقاوم AEC به مدت طولانی توسط L-لیزین مهار نمی شوند و منجر به افزایش بازده لیزین می گردند .
از زمان شناسایی توالی ژنومی C.glutamicum در سال 2003 , و تمامی ژن ها برای انزیم های کلون شده در گیر در این مسیر کد شدند . روش های ژنتیکی و مهندسی متابولیک بر پایه تجزیه و تحلیل شار نقش مهمی را حتی در دستیابی به سویه های جهش یافته قوی ایفا می کنند . امروزه , بازدهی در محدوده > 100 g L–1 در 60 ساعت , و (به عنوان مثال برای L-گلوتامات ) تکرار پروتکل های fed-batch در بیوراکتورها تا سقف حجم 75 متر مکعب استفاده می شود .
منبع کربن معمولا از شیره نیشکر , و تجزیه سوبسترا 75 گرم L-لیزین را در هر 100 گرم گلوکز میتوان بدست اورد .
محتوای بیوتین در محیط کشت باید بیشتر از 30 میلی گرم در لیتر باشد . بازیابی ممکن است به وسیله اسپری خشک کننده (spray-drying ) و گرانولیزاسیون کل به وجود آمده ادامه یابد و یا از مسیر جداسازی سلول ها با روش کروماتوگرافی تبادل یونی و کریستاله کردن انجام پذیرد . یک جایگزین مناسب , هر چند از لحاظ اقتصادی قابل قبول و دوام نیست , تولید D,L-α-amino-ε-caprolactam (ACL) به عنوان یک حد واسط ارزان از سنتز شیمیایی Nylon™ , با سلول های خشک شده استون از Cryptococcus laurentii در یک راکتور سلولی می باشد .
L-ترئونین :
ارگانیسم های انتخاب مسیر تولید این آمینواسید جهش های E.coli را رد میکنند . اپرون ترئونین کلون شده است و برای بهبود بیشتر سویه ها استفاده می شود . بازیابی توسط جداسازی سلول , به روش اولترا فیلتراسیون وبعد کریستالیزاسیون محصول حاصل از فیلتراسیون آغاز می شود .
جنبه اقتصادی لیزین ، متیونین و ترئونین :
تولید سالانه L- لیزین در سال 2009 در حدود 3/1 میلیارد تن , LوD –متیونین حدود 850000 تن و L-متیونین حدود 190000 تن بوده است . فرآیند مورد نظر برای تولید متیونین سنتز شیمیایی راسمات (racemate ) است. L -ترئونین و L-لیزین به صورت انحصاری به روش تخمیر تولید می شوند. قیمت هر یک تن از این سه اسید آمینه بین1500 تا 4000 دلار امریکا است. که بازاری به ارزش معادل 3 بیلیون دلار امریکا دارد .
در آینده دوررتر , انتظار می رود که اسیدآمینه های تولید شده به روش صنعتی به محصولات خوراکی اضافه شوند و رقابت ناشی از نسل محصولات تراریخته (transgenic) حاوی ترکیبات بهینه سازی شده آمینواسید برای اهداف تغذیه ای حاصل شود.
نویسنده : طاهره کیااحمدی – کارشناسی میکروبیولوژی
